
Written by Peter Joffe
In this article, I will share my thoughts on using blood lactate testing for training athletes.
For over a century, lactate has been strongly linked to and even blamed for fatigue.
Therefore, it should come as no surprise that the word “lactate” is often used in the sports community.
It is often referred to support certain strict and “science based” prescriptions.
However, do we have a clear understanding of all aspects of lactate metabolism and its role in exercise?
Lactate measurements are traditionally used for exploration of endurance capacity and for attempts to locate Anaerobic Threshold (AT) . So it is not a coincidence that other names for AT are connected with lactate. Lactate Threshold, Lactate Turn Point, Maximal Lactate Steady State, Minimum Lactate Equivalent, Onset of Blood Lactate Accumulation, you name it.
In the past, BL testing was possible only in laboratories.
Now portable lactate meters are commercially available. They are quite reliable and simple to use. However, BL measurement is invasive, carries a risk of infection, and may be unpleasant for some people.
I’m ready to bleed for science, but when dealing with athletes, your scientific curiosity should not distract the training process. It is why you have to weigh the pros and cons.
“Far from being a dead-end metabolite, Lactate stands at the crossroads of metabolism”. Gladden, L.B., “A “Lactatic” Perspective on Metabolism”.
Why lactate?
When developing endurance, we are particularly interested in intensity ranges from AT to maximum. Basically, in this range, most elite athletic events occur, and it is crucial for many sports games and combat sports.
What is the anaerobic threshold, is the topic which continues to be debated for many years. For this article, I will stick with two possible meanings:
1. Intensity above which energy demand cannot be met by exclusively aerobic production.
2. Highest intensity where relatively long stability of homeostasis is possible.
Exploring energy supply above AT we are interested mostly in carbohydrates (glucose/glycogen) metabolism. It is because other main energy sources such as fat and protein need oxygen for utilisation (so they are aerobic). “Pure” anaerobic reserves such as Creatine Phosphate (CrP) and intracellular ATP can fuel exercise only for a few seconds. Carbohydrates can do it for tens of minutes.
Differently from fat, protein, and CrP, which are mono-pathway fuel, carbohydrates can be used both aerobically and anaerobically. Thus perhaps we can consider their transition between two pathways as a basis for AT definition.
Traditional concept.
The next paragraph describes how we were taught in universities several decades ago, and such understanding is still the mainstream not only for sports practitioners but also for academia.
The first stage of glucose/glycogen metabolism does not require oxygen and produces two net ATP on the way to conversion to pyruvate. Then, if oxygen is available, pyruvate goes for oxidation in mitochondria. The end products of this process are CO2, H2O, and, importantly, around 30 ATP.
If there is insufficient oxygen supply, part of pyruvate is converted to lactic acid. That allows to maintain redox status and to continue glycolysis. Nevertheless, lactate, as a waste product, causes acidosis and eventually work termination. So appearance lactate in blood is a manifestation of anaerobic energy production and lack of oxygen (Picture 1).
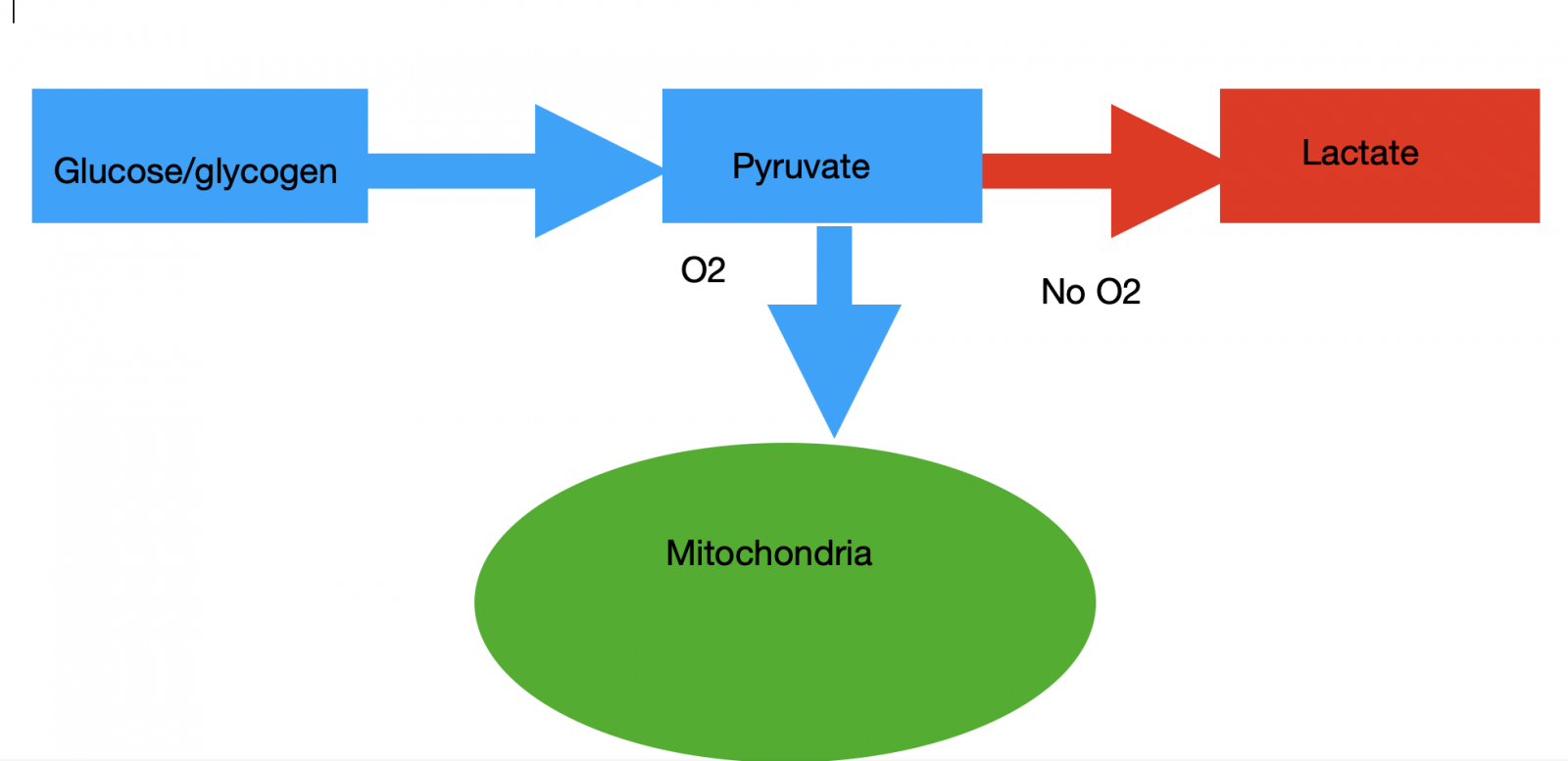
Picture 1. Traditional view on carbohydrates metabolism.
Something is wrong.
However, a few interesting facts do not fit this concept.
1. Lactate is present in the blood even at rest (O2 is more than enough).
2. Exponential accumulation of lactate occurs at intensities lower than those associated with VO2 max; thus, the oxidative capacity is not exhausted yet.
3. Highlanders, Sherpas and Andean people, exhibit the so-called “lactate paradox”. When they do exercise of a given intensity at a higher altitude they produce less lactate. So, under more and more hypoxic conditions they produce less and less lactate (Hochachka, 2018). That is strange, isn’t it?
4. Hypoxia does not result in less VO2 for given sub-maximal work though it results in less VO2 max.
I’ll try to explain this point.
If you are doing VO2 max test at sea level and at 4000 m, yes, at altitude your VO2 max and work rate at VO2 max will be lower. However, if you do some sub-maximal work, which does not elicit VO2 max, you will achieve the same VO2 for the same intensity regardless of altitude.
Thus, within the certain intensity range, the body can adjust complex oxidative metabolism and maintain the same rate of aerobic energy production even in hypoxia. However, BL often rises in these conditions, and this is unclear why. If VO2 is the same, and work/intensity is the same: what additional “hypoxic BL” paid for? (Picture 2).

Picture 2. Additional lactate in hypoxia.
The same sub-maximal work at normoxia and hypoxia (red+blue rectangles) results in the same VO2 (blue rectangle). However, additional lactate is formed in hypoxia (brown rectangle). If this lactate was the result of an increase in anaerobic contribution, VO2 would have to decrease.
Another idea of lactate metabolism during exercise.
The new concept of lactate metabolism suggests that lactate formation goes continuously even in fully aerobic conditions. Even more, some scientists argue that actually formation of lactate is the main pathway of glycolysis in any condition. Even at rest, lactate to pyruvate ratio is 10:1 and goes up to 160:1 at moderate exercise (Brooks, 2020).
So there is no such thing as aerobic and anaerobic glycolysis. There is only one glycolysis which ends up with lactate formation (Schurr, 2017).
It is what happened next can be seen as a fork of the aerobic and anaerobic roads. If an organism can utilise lactate aerobically it is combusted or converted into glucose in gluconeogenesis. If not, it starts to accumulate in working muscles and eventually in blood (Picture 3).

Picture 3. Lactate stands on a cross-road of metabolism.
Lactate and Anaerobic Threshold.
Due to the balance between production and utilisation, the BL concentration is stable up to some intensity, although this stability can be at different levels. Usually, at higher intensities, the stabilisation level is higher. For example, at rest, BL can be stable at 1 mmol / l, with moderate exercise 2 mmol /l, and with heavy exercise more than 4 mmol /l.
It is well known that the heart, kidneys, brain, and liver willingly consume lactate as fuel or substrate for gluconeogenesis. Resting and working muscles are the significant consumers of lactate as well. So, many organs can take-up lactate. However, there is one important point:
Oxygen is essential for all pathways for lactate utilisation. Thus, all lactate clearance is, in essence, aerobic.
Thus, we came to an interesting conclusion: since the utilisation of lactate can only be aerobic, and the accumulation of lactate is the only way for “long-term” anaerobic energy production (longer than a few seconds), we can say that if BL is stable, all energy is produced aerobically (Peronnet, 2010).
And only when lactate starts to rise consistently at the constant intensity we may assume that aerobic capacity cannot cope with energy demand and is supplemented by anaerobic energy production.
This moment and intensity can be called anaerobic threshold if you wish.
It is the accumulation of lactate or other glycolytic intermediates, not simply evidence of lactate production, which should be considered to represent the metabolic rate above the anaerobic threshold (Svedahl, 2003).
Lactate and oxygen.
Relation lactate with oxygen may be more complex than we previously thought.
As I already said, there are situations when oxidative capacity is still available; however, BL starts to build up. In some circumstances, hypoxia even leads to decreasing lactate concentration—“lactate paradox”. Does this mean that lactate accumulation is independent of oxygen availability? No, this claim is not correct.
Mitochondria power is oxygen-dependent but not oxygen limited.
On the contrary, it is a well-known fact that the availability of oxygen affects the formation of lactate in most cases. However, now we understand that it is perhaps indirectly and that oxygen is not the only factor.
What is a possible explanation?
Oxygen and lactate may be connected through the cell’s energetic status — ATP/ADP;Pi ratios. Oxygen supply has some inertia and increasing intensity results in its temporary deficit, which impairs oxidative ATP production and decreases ATP/ADP;Pi ratio (Ferguson, 2018). That stimulates both mitochondria, which “step-up” oxidative phosphorylation and glycolysis, consequently lactate formation. ATP production increases, therefore energetic status, and BL may stabilise at a new level. BL will be higher, whereas energetic status— lower (Peronnet, 2010).
In a wide range of limiting oxygen availability, mitochondria still can maintain their “inner” VO2 consumption at sub-maximal exercise. However, this adjustment includes a mechanism that leads to increased glycolysis, and therefore increased lactate production.
Thus, usually, in hypoxic conditions arising from altitude or exercise, BL concentration may be higher. But, as was said, if it’s stable, there are no significant anaerobic energy production (Picture 4).

Picture 4. Lactate in hypoxia.
Higher blood lactate not necessary means increased anaerobic contribution. In fact, if BL is stable, it is completely utilised in oxidative metabolism.
Lactate paradox.
This dependence of BL concentration on ATP/ADP; Pi ratio may help us to understand the “lactate paradox”.
Highlanders have better regulation of ATP demand-production coupling. For subjects accustomed to oxygen deficit due to long-term altitude exposure, smaller ATP/ADP ratio changes are sufficient for mitochondria to increase oxidation rate. This allows highlanders to avoid dramatic perturbations in the cell’s energetic status (Hochachka, 2018).
Highlanders exercising higher become more economical and decrease production of glycolytic ATP. The rate of glycolysis is reduced and hence the production of lactate. Though decreasing energy production results in lower VO2max, highlanders are still much more productive than non-accustomed lowlanders, perhaps due to less metabolites accumulation.
By the way, endurance athletes may possess similar mechanism. They usually produce less lactate at a given intensity than their non-endurance colleagues as well. Though, of course, many other factors are involved.
They will be discussed in the next paragraph.
Why AT is different from VO2 max?
Another interesting question is why the exponential build-up of BL happens before VO2 max is reached?
Basically, this means that aerobic capacity is still available; however, the rate of lactate production outstrips its consumption rate. Why can not the body increase the rate of lactate utilisation and maintain its stable level in blood?
The problem may be in transportation.
Lactate is produced in one place and eventually utilised in another, so it needs to be transported there.
To explain that, George Brooks three decades ago proposed a “Lactate shuttle theory” (Brooks, 2018).
It is widely accepted now that lactate shuttles transport lactate from production cells to the cells of utilisation. This is called a cell-to-cell shuttle. Lactate consumers are the liver, heart, brain, etc. With increasing intensity, muscles start to play a major role. Lactate travels from working muscles to resting ones and from glycolytic fibres to oxidative.
All these need good transportation network. Blood flow rate, capillary density, capillaries-per-fibre ratio, and shuttles capacity play a major role.
Lactate can be utilised within the muscle cell as well. For that, it should be transported from cytosol to mitochondria or its vicinities. How this happens exactly is still debated (Gladden, 2007) but it looks proven that intercellular lactate consumption exists (Gladden, 2008).
In this sense, oxidative slow-twitch fibres may be really crucial. They are not only high-utiliser of external lactate from fast-twitch fibres but also high-consumer of their own lactate.
From this point of view, the ability of elite endurance athletes to maintain stable lactate at intensities close to VO2 max may be explained by:
1. High percentage of slow-twitch fibres (lower lactate production and it oxidation “on the spot”).
(Ivy, 1980.)
2. High capillary density ( good transportation network).
(Joyner, 2008)
3. High mitochondria volume density (more lactate utilisation “factories” are available).
4. Fine regulation of demand-production coupling (lower metabolites production).
Less-trained or non-endurance athletes have LT further from VO2 max; thus significant amount of ATP starts to produce anaerobically long before aerobic capacity is exhausted. This results in faster glycogen depletion and faster build-up of lactate and exercise byproducts.
Conclusion for Lactate – Oxygen:
‘Glycolysis resulting in the formation of lactic acid should be interpreted as a process occurring without the use of oxygen, not necessarily in the absence of oxygen. It is now recognised that although hypoxia may result in increased formation of lactic acid, the absence of oxygen is not a prerequisite for lactic acid formation” (Svedahl, 2003).
Lactate and fatigue.
Why blood lactate testing is so important in the first place?
Lactate accumulation is perhaps a direct indicator that part of the energy is produced anaerobically. The rate of build-up tells us exactly how dramatically aerobic capacity is exceeded.
More importantly, for practical coaching, lactate may give us valuable information about a disturbance in homeostasis.
For many years lactate was blamed for such disturbances. It was considered a direct cause of acidosis. Lactic acid dissociates with production of H+ ions. H+, in its turn, inhibits energy production (Conley, 2001). Now we understand that it is probably not so simple. Lactate reflects the accumulation of H+, but it is debatable if it produces them. Without a doubt, glycolysis creates metabolites, including H+, but things will probably be much worse without lactate. Some authors argue that lactate, in fact, buffers H+ and facilitate its removal (Robergs, 2004). Definitely, fighting with lactate does not prevent acidosis.
In conclusion: Acidosis is a consequence of insufficient mitochondrial oxidation, and the same is BL accumulation (Conley, 2001). So these two are pretty well correlated. However, correlation does not mean cause-effect.
An alarm signal.
So most likely lactate can not be blamed for disturbance in homeostasis. At least as a direct and the main cause of fatigue.
However, it may be a useful indirect indicator of it.
The logic is simple: If an organism for any reason cannot utilise lactate, it probably cannot utilise other metabolites as well. For example, H+, and Pi.
However, it is important to understand that the word “indirect” is crucial. There is no particular level of lactate which manifests work termination. Sometimes athletes can still exercise relatively long at 10 mmol/l, but sometimes terminate at 3 mmol/l. People can definitely be fatigued with different BL and without lactate at all (e.g. McArdle disease).
For example, (Jones, 2006) and (Coyle, 2005) reported exceptionally low BL levels after maximum exercise in two superstar endurance athletes. Their BL after VO2 max test was lower than normal values even for top athletes. Unfortunately, we still don’t know exactly why. And even with these low BL levels, they still were fatigued.
Our own research at the University of Exeter showed no correlation between performance and lactate at fatigue in relatively long exhaustive exercise (BL test). At the same time, lactate curves were more informative (Picture 5).
So, you need to look not only at the specific BL values, but the whole picture.
Lactate dynamic, which reflects the rate of glycolysis , oxidative capacity, transportation capacity, and buffering capacity, may give us useful information about overall athlete’s endurance and its type.


Picture 5. The results of blood lactate testing for six subjects.
Upper panel: no correlation between end-blood lactate and performance in six subjects. Lower panel: right-shift in BL curves indicates (albeit not perfectly ) better performance.
Key points before using BL in practice.
So what can we can take on board from the analysis above before doing practical blood lactate testing?
1. The anaerobic pathway can be identified as a consistent accumulation of lactate, not merely by its presence. Thus one measurement which resulted in even as high BL as 10 mmol/l does not necessarily tell us that there is significant anaerobic production. We need to look at dynamics.
2. During exercise, lactate concentration indirectly indicates the level of perturbation in cells. That is the cell’s energetic and redox state. Thus higher lactate during exercise, while not necessarily mean anaerobic contribution, may indicate that the athlete is exerting more effort to maintain “aerobic status.”
3. Lactate disposal to a great extend depends on mitochondria capacity and the body’s transportation network (capillaries and shuttles) . The same is important for other metabolites utilisation. Thus stable lactate is a good indirect indicator of overall homeostasis sustainability.
Blood lactate testing.
Standard tests.
Usually, blood lactate testing aims to find AT.
MLSS (Maximal Lactate Steady State) test is widely considered as a Gold Standard for that (Billat, 2003). That follows straightforwardly from the notion that maximal intensity where lactate stability is possible is a border after which anaerobic production begins. That, in essence, is correct.
However, there are some methodological problems with this test.
For example, Jones argues that excepting MLSS as a Gold Standard for anaerobic threshold is questionable (Jones, 2019).
He asks, quite correctly: Where criteria for this test, such as the rise of BL for 1 mmol/l or taking samples exactly in 10 and 30 min, came from? They are quite arbitrary and are not based on scientific rationale. I totally agree.
Additionally, MLSS test is not practical. It demands 3-5 30 min efforts on separate days.
Another common way of blood lactate testing is the standard BL test.
It consists of 8-10 3 min stages with stepwise increasing intensity. The treadmill provides a more controlled environment for the running, whereas a field test is more ecologically valid.
Usually, 1 km/h increment is used, which gives an additional 50 m distance for each new 3 min stage in the field test. BL is measured at the end of each stage. Then BL is plotted against intensity/speed, which produces BL curve.
Again, the aim is to find AT (LT, LTP, etc.).
However, how can we choose some particular, “true” threshold point on a curve?
The curve has no thresholds, and all methods of setting some point on it as threshold are subjective assumptions or mathematical manipulations.
I described math behind one of the most popular methods— D-max in the article and made a calculator for it automatic determination. There is no physiology behind it, just math. So, let me be clear: BL curve has a physiological basis. Choosing some point on it as a lactate threshold does not (Tokmakidis, S.P., Léger, L.A. and Pilianidis, T.C., 1998).
There are two other modifications of standard blood lactate testing— Individual Anaerobic Threshold (IAT) and Minimal Lactate Equivalent (MLE).
Both of them again aim to find AT.
The first include math and/or subjectivity. In second, AT intensity is derived from physiological measurement but its validity and reliability is questionable.
For reader interested in MLE more:
Arguments pro: (Garcia, 2018) and con: (Carter, 1999).
Hunting of the snark.
The key question for me regarding blood lactate testing is:
Why do we keep looking for AT in BL testing? Why do we think that some intensity found in artificial conditions would be so important? Is there a unique AT intensity at all?
Indeed, different tests, different test designs and AT definition methods result in different AT intensities (Faude, 2009).
Still, scientists continue to hunt for some Gold Standard anaerobic intensity.
For example, Jones, opposing the universal value of the MLSS test, proposed another test for the role of the Gold Standard — Critical Power (Jones et al. 2019), which, in my opinion, is no less dubious than MLSS (see article).
From my point of view, there is no Gold Standard Test for AT because there is no such Gold Standard AT (good analysis in: Hopker, 2011). What really exists is a state of homeostasis when exercise becomes progressively difficult, and fatigue looms on the horizon. That may happen at different intensity and different times depending on many interacting factors.
Regarding to threshold speed, for many years, athletic coaches use reference velocities (e.g. over 10000 m) for designing “threshold runs”. They often do not use blood lactate testing, though they may refer lactate as a scientific basis for their prescriptions. And they are successful!
We may use some point on the BL curve as reference for AT, especially for athletes who are not runners and do not know so precisely their speeds on different distances. However, this chosen speed will not be some magic threshold and should be adjusted empirically.
“Several tests have been developed to determine the intensity of exercise associated with AT: maximal lactate steady state, lactate minimum test, lactate threshold, OBLA, individual anaerobic threshold, and ventilatory threshold. Each approach permits an estimate of the intensity of exercise associated with AT, but also has consistent and predictable error depending on protocol and the criteria used to identify the appropriate intensity of exercise. These tests are valuable, but when used to predict AT, the term that describes the approach taken should be used to refer to the intensity that has been identified, rather than to refer to this intensity as the AT” ( Svedahl, K. and MacIntosh, B.R., 2003).
Curve position.
In my opinion, the BL curve itself gives us much more valuable information than just a subjective definition of the AT. For that we may analyse its position against other parameters.
BL-intensity.
The right shift of the curve on the speed/intensity scale means improving fitness.
For example, Jones showed how the shift of the BL curve indicated significant progress for Paula Radcliffe. over the years leading up to her world record (.Jones, 2006).

Picture 6. Paula Radcliffe BL curve right shift over ten years.
Adopted from ((Jones, A.M., 2006)
If the athlete moved BL curve right against speed (Picture 6) it is good, but it is difficult to answer what exactly improved. It may be some “biomechanical ” improvements. I mean improved technique, strength, lean mass and even shoes. Or that may be “biochemical” enhancements (VO2 max, metabolite clearance, etc.)
For example, in the work mentioned, Jones pointed out that Paula Radcliffe. has become a much more economical runner in terms of oxygen consumption, despite the same VO2 max. One of the variable that improved was biomechanical— countermovement jump.
Can we somehow to separate biomechanical and biochemical factors? It would be interesting to analyse this question a little bit further.
Thus another way of plotting BL may be useful.
BL—% of VO2 max.
That is plotting BL against % of VO2 max. The shifted BL curve on %VO2 max scale tells us that athletes can perhaps sustain higher VO2 max fraction for longer, which is a really important quality. (Joyner, 2008) .
Indeed, the VO2 max indicates oxidative power. BL curve indicates a disturbance in homeostasis.
If BL shifts right on the %VO2 scale, we may rule out exclusively biomechanical factors and even VO2 max itself. Otherwise, the fractional VO2 consumption remained the same despite the improved performance (Picture 7).

Picture 7. Hypothetical cyclist moved his AT from 260 to 290 W: How he paid for that?
Initial position of BL curve (blue line). AT happened at 70% of VO2 max (variant 0). Athlete improved —curve shifted right (red line). This may be due to improved economy (paid the same VO2 for higher intensity , variant 1). Or due to improved VO2 max (paid more VO2 in absolute numbers but the same 70% fraction, variant 2). Or due to larger fractional utilisation ( utilised 80% of VO2 max at AT).
Thus moving right on % VO2 max scale means that the athlete starts working closer to maximal oxidative capacity while still maintaining stable homeostasis . That perhaps reflects “metabolical improvements,” for example, capillarisation (Joyner, 2008).
This high fractional utilisation is a trait of top-class endurance athletes. They have AT ( dependent on the intensity you choose for it) at 80-90 % of VO2 max. It is a great advantage in a wide spectre of endurance events.
Of course, you need a laboratory for testing together BL and VO2.
However, do not expect that such parameter as a fractional utilisation of VO2 max improves fast. It may take years. Thus booking laboratory for blood lactate-VO2 testing once-twice a year may be more than enough.
BL—HR.
Plotting BL against HR or HRR (heart rate reserve) gives you some general information about HR parameters at particular point on BL curve.
For example, in every fitness device you have option of monitoring “intensity zones” based on % of HR max. Usually there are five of them: 50-60 % of HRmax; 60-70; 70-80; 80-90 and 90-100. There may be slightly different divisions but you have got the idea.
Of course, this is too general. Starting from the fact that they used your predicted maximum heart rate based on the formula (220 – age), which is not always true, and ending with the assumption that the same% heart rate values are suitable for everyone.
So connecting HR with individual BL curve gives at least some physiological basis for HR-based intensity zones.
For instance, you may choose HR or HRR before first rise of lactate as zone 1, HR half-way between first rise and exponential rise as Zone 2, from that until exponential rise as Zone 3 and so on (Picture 8).

8. HR and blood lactate for 38 years old subject during incremental cycle test.
HR may be assigned as: Zone 1 – before first rise in BL; Zone 2 – between first rise and half-way to exponential rise; Zone 3 between half-way and exponential rise; Zone 4 after exponential rise and halfway to maximum; Zone 5 – from half-way to maximum
However, it is necessary to note that the usefulness of this approach may be limited by HR fluctuations, independent of factors affecting the BL curve. Additionally, relationship between BL and HR found in standard test is not always applicable to other mode of exercise. Nevertheless, it is probably better than fitness watches guidance anyway.
In fact, with the training, both HR and BL graphs moved right on intensity scale . For example, Jones reported that Paula Radcliffe HR at LT remains the same for ten years (180 b/min). However, this parameter shifted right from 14 km/h to a remarkable 18.5 km/h in that period (Jones, A.M., 2006; Picture 9).

Picture 9. Paula Radcliffe HR right shift over ten years.
Adopted from ((Jones, A.M., 2006)
In conclusion, plotting BL against HR/HRR and both parameters against intensity is not difficult (anyway, athletes often run with HR monitors). That may give you additional information about course of adaptations.
Conclusion for standard blood lactate testing:
Some intensity, arbitrarily assigned as AT after testing BL, may be beneficial, but has no real advantage over other empirical approaches used by sports trainers for many years.
However, BL curve itself may provide useful information about an athlete’s endurance capacity.
So maybe stop just hunting for the lactate threshold, and to look at the lactate curve more comprehensively?
Monitoring training.
Buffering
Achieving a high BL is beneficial for a variety of maximal workouts from 10 seconds to 1 minute. In this type of exercise, aerobic capacity is clearly not enough to prevent the rapid accumulation of lactate (and other metabolites), and this is exactly what we want. We aim to develop the ability to maintain maximum intensity for longer. Physiologically this ability is called “buffering”. Coaches may call it “speed endurance”.
Usually, researchers are bothered with buffering acidosis, maybe because we are relatively good at measuring it. I suppose there are other metabolites, for example, Pi (inorganic phosphate), which is the product of ATP hydrolysis. I am not competent enough to go deeper on buffering. In practice, it is important that in speed endurance exercises Bl is a reliable indicator of anaerobic production. Since many metabolites are either produced anaerobically or utilised aerobically, a high BL means a high contamination rate. And the ability to maintain intensity in these conditions is sometimes really important.
Lacour et al. found a strong positive correlation between 400 m performance and BL level after race among elite runners (Lacour, 1990). Thus ability to produce energy anaerobically and tolerate high level of by-products is crucial for this distance.
Interestingly, BL was almost the same after 400, 800, and 1500 m. That possibly indicates that in distances lasting 1-4 min, anaerobic production and buffering capacity still has to be extremely high. However, the correlation between BL and performance in 800 and 1500 m was less than in 400 m, probably due to increasing aerobic influence.
So it looks like for this group of elite runners, extremely high BL in the range of 22-27 mmol/l indicates remarkable buffering capacity. This limit can be rapidly achieved in 400 m (46 sec) or can be stretch due to pacing for 1:45 (800) and 3:35 (1500). The basic idea is that high lactate level is good for training metabolite tolerance, and you can peak during an effort of less than one minute.
Decreasing BL without compromising performance in such workouts indicates adaptation. Training stimulus maybe not enough any more, and the coach may consider changing something. Perhaps it would be useful to do blood lactate testing quite often on this type of training, but the coach has to take into account practical issues.
Sub-maximal exercise.
Another implication of blood lactate testing in training may be looking at lactate during intense but sub-maximal exercise (between AT and VO2 max).
Theoretically, lower BL in such efforts may be an indicator of better overall fitness and/or current form (Coyle, 2005). In my opinion, it is better to compare athletes against themselves than against others. BL values have significant interpersonal variability and inside a homogeneous group, BL may not be a reliable indicator of fitness. For example, (Myburgh et al, 2001) found no correlation between BL and performance in one-hour race for cyclists.
I don’t think it worth measuring BL often in sub-maximal exercise, but sometimes you can. For example, a combination of lower BL, RPE (Rate of Perceived Exertion) , and HR while running at a given sub-maximal speed is a reliable indicator of improved aerobic fitness.
BL recovery.
The idea to measure BL recovery is based on the assumption that when lactate production stops, speed of recovery indicates clearance capacity. The faster—the better.
However, recovery depends on many factors, including previous workouts, recovery mode, and the current form.
Thus, in my opinion, too many factors are involved in BL recovery, so it implementation in training is impractical. If you want to use BL recovery rate in a more controlled way, there is an IAT test, which uses the rate of recovery for AT definition.
Conclusion.
Without a doubt, lactate is an important product of human metabolism in all conditions, including exercise.
Although it has been studied for many years, its exact role and functions are still being discovered (Brooks, 2020; Gladden, 2004).
In sports, blood lactate testing are most commonly used to chase AT.
Well, blood lactate testing can be used for definition of some training intensity, which can be used for “threshold training,” but we have to understand that it is just one of the possible approaches, and such intensity is not a universal and unique value.
Exercise fatigue is too complex and multifactorial to be described by any simple boundary.
It can and should be investigated by various methods, including BL measurements.
Definitely, BL can be really good at this task because it plays a crucial role in exercise metabolism. We have just begun to understand and appreciate that. From this point of view, lactate, which was once considered our nemesis, is in fact our valuable friend.
References.
Acevedo, E.O. and Goldfarb, A.H., 1989. Increased training intensity effects on plasma lactate, ventilatory threshold, and endurance. Medicine and science in sports and exercise, 21(5), pp.563-568.
Beneke, R., 1995. Anaerobic threshold, individual anaerobic threshold, and maximal lactate steady state in rowing. Medicine & science in sports & exercise, 27(6), pp.863-867
Billat, V.L., Sirvent, P., Py, G., Koralsztein, J.P. and Mercier, J., 2003. The concept of maximal lactate steady state. Sports medicine, 33(6), pp.407-426.
Brooks, G.A., 2020. Lactate as a fulcrum of metabolism. Redox biology, 35, p.101454.
Brooks, G.A., 2018. The science and translation of lactate shuttle theory. Cell metabolism, 27(4), pp.757-785.
Cairns, S.P., 2006. Lactic acid and exercise performance. Sports medicine, 36(4), pp.279-291.
Carter, H.E.L.E.N., Jones, A.M. and Doust, J.H., 1999. Effect of incremental test protocol on the lactate minimum speed. Medicine and Science in Sports and Exercise, 31(6), pp.837-845.
Conley, K.E., Kemper, W.F. and Crowther, G.J., 2001. Limits to sustainable muscle performance: interaction between glycolysis and oxidative phosphorylation. Journal of Experimental Biology, 204(18), pp.3189-3194.
Connett, R.J., Honig, C.R., Gayeski, T.E.J. and Brooks, G.A., 1990. Defining hypoxia: a systems view of VO2, glycolysis, energetics, and intracellular PO2. Journal of Applied Physiology, 68(3), pp.833-842.
Coyle, E.F., 2005. Improved muscular efficiency displayed as Tour de France champion matures. Journal of Applied Physiology.
Coyle, E.F., Martin 3rd, W.H., Sinacore, D.R., Joyner, M.J., Hagberg, J.M. and Holloszy, J.O., 1984. Time course of loss of adaptations after stopping prolonged intense endurance training. Journal of Applied Physiology, 57(6), pp.1857-1864.
Faude O, Kindermann W, Meyer T.,2009. Lactate threshold concepts: how valid are they?. Sports Med. 2009;39(6):469-490. doi:10.2165/00007256-200939060-00003.
.Ferguson, B.S., Rogatzki, M.J., Goodwin, M.L., Kane, D.A., Rightmire, Z. and Gladden, L.B., 2018. Lactate metabolism: historical context, prior misinterpretations, and current understanding. European journal of applied physiology, 118(4), pp.691-728.
Garcia-Tabar, I. and Gorostiaga, E.M., 2018. A “blood relationship” between the overlooked minimum lactate equivalent and maximal lactate steady state in trained runners. Back to the old days?. Frontiers in physiology, 9, p.1034.
Gladden, L.B., 2008. A” lactatic” perspective on metabolism. Medicine & Science in Sports & Exercise, 40(3), pp.477-485.
Gladden, L.B., 2004. Lactate metabolism: a new paradigm for the third millennium. The Journal of physiology, 558(1), pp.5-30.
Gladden, L.B., 2007. Is there an intracellular lactate shuttle in skeletal muscle?. The Journal of physiology, 582(Pt 3), p.899.
Hochachka, P.W., Beatty, C.L., Burelle, Y., Trump, M.E., McKenzie, D.C. and Matheson, G.O., 2018. The lactate paradox in human high-altitude physiological performance. Physiology.
Hopker, J.G., Jobson, S.A. and Pandit, J.J., 2011. Controversies in the physiological basis of the ‘anaerobic threshold’ and their implications for clinical cardiopulmonary exercise testing. Anaesthesia, 66(2), pp.111-123.
Ivy, J.L., Withers, R.T., Van Handel, P.J., Elger, D.H. and Costill, D.L., 1980. Muscle respiratory capacity and fiber type as determinants of the lactate threshold. Journal of Applied Physiology, 48(3), pp.523-527.
Jones, A.M., 2006. The physiology of the world record holder for the women’s marathon. International Journal of Sports Science & Coaching, 1(2), pp.101-116.
Jones, A.M., Burnley, M., Black, M.I., Poole, D.C. and Vanhatalo, A., 2019. The maximal metabolic steady state: redefining the ‘gold standard’. Physiological reports, 7(10), p.e14098.
Joyner, M.J. and Coyle, E.F., 2008. Endurance exercise performance: the physiology of champions. The Journal of physiology, 586(1), pp.35-44.
Lacour, J.R., Bouvat, E. and Barthelemy, J.C., 1990. Post-competition blood lactate concentrations as indicators of anaerobic energy expenditure during 400-m and 800-m races. European journal of applied physiology and occupational physiology, 61(3-4), pp.172-176.
MYBURGH, K.H., VILJOEN, A. and TEREBLANCHE, S., 2001. Plasma lactate concentrations for self-selected maximal effort lasting 1 h. Medicine & Science in Sports & Exercise, 33(1), pp.152-156.
Nalbandian, M. and Takeda, M., 2016. Lactate as a signaling molecule that regulates exercise-induced adaptations. Biology, 5(4), p.38.
Peronnet, F., 2010. Lactate as an end-product and fuel. Deutsche Zeitschrift fur Sportmedizin, 61(5), p.112.
Robergs, R.A., Ghiasvand, F. and Parker, D., 2004. Biochemistry of exercise-induced metabolic acidosis. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 287(3), pp.R502-R516.
Svedahl, K. and MacIntosh, B.R., 2003. Anaerobic threshold: the concept and methods of measurement. Canadian journal of applied physiology, 28(2), pp.299-323.
Schurr, A., 2017. Lactate, not pyruvate, is the end product of glucose metabolism via glycolysis. Carbohydrate.
Tokmakidis, S.P., Léger, L.A. and Pilianidis, T.C., 1998. Failure to obtain a unique threshold on the blood lactate concentration curve during exercise. European journal of applied physiology and occupational physiology, 77(4), pp.333-342.